全國服務(wù)咨詢熱線:
15221734409
15221734409

n 研究背景
卵泡刺激素(FSH)是驅(qū)動卵泡發(fā)育的關(guān)鍵內(nèi)分泌信號,雖已知其能促進(jìn)顆粒細(xì)胞(GCs)的糖酵解和乳酸生成,但糖酵解通量如何與線粒體生物發(fā)生相偶聯(lián)的機(jī)制尚不清楚。近年來,乳酸被重新認(rèn)識為一種生物活性代謝物,可通過組蛋白乳酸化修飾將細(xì)胞代謝狀態(tài)與染色質(zhì)結(jié)構(gòu)和基因調(diào)控聯(lián)系起來。然而,這種代謝-表觀遺傳調(diào)控機(jī)制是否在FSH介導(dǎo)的卵泡發(fā)育生理過程中發(fā)揮作用,特別是如何協(xié)調(diào)線粒體生物發(fā)生以滿足卵泡生長的能量需求,仍是未解之謎。

n 研究結(jié)果
2.1 FSH誘導(dǎo)的線粒體生物發(fā)生伴隨卵巢顆粒細(xì)胞中H4K5la升高
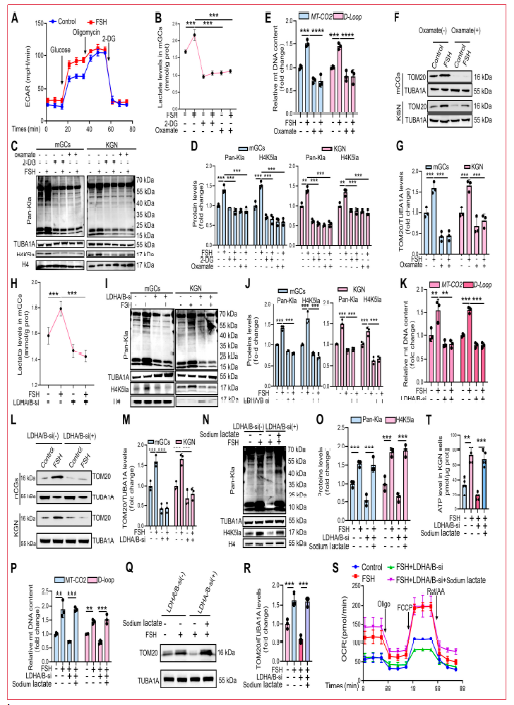
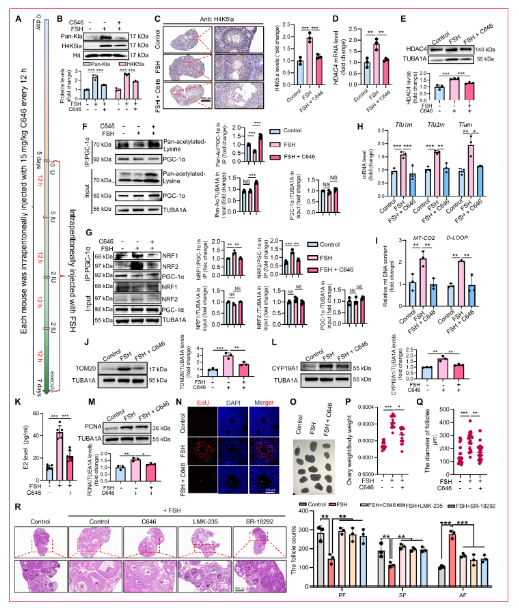
研究人員通過腹腔注射FSH評估其對小鼠卵巢顆粒細(xì)胞(mGCs)線粒體生物發(fā)生的影響。結(jié)果顯示,F(xiàn)SH刺激增加了mGCs中線粒體DNA編碼的細(xì)胞se素c氧化酶II(MT-CO2)基因和mtDNA D-loop區(qū)域的拷貝數(shù),線粒體標(biāo)志物TOM20水平升高。透射電鏡進(jìn)一步顯示FSH顯著增加mGCs中線粒體數(shù)量,同時保持超微結(jié)構(gòu)完整性,包括膜形態(tài)和嵴組織,線粒體體積也增加,提示FSH暴露后線粒體融合增強(qiáng)。FSH處理還顯著上調(diào)線粒體融合蛋白MFN1、MFN2和OPA1表達(dá),而裂變蛋白DRP1及其Ser616磷酸化水平降低,Ser637磷酸化增加,表明線粒體裂變受抑制。FSH處理還顯著提高了mGCs中ATP水平。由于FSH在卵泡發(fā)育過程中增加糖酵解導(dǎo)致乳酸升高,研究人員評估了乳酸是否促進(jìn)FSH介導(dǎo)的線粒體生物發(fā)生。FSH注射后,mGCs表現(xiàn)出顯著增加的葡萄糖轉(zhuǎn)運蛋白1(GLUT1)表達(dá)和乳酸水平。免疫組織化學(xué)顯示FSH處理后卵巢組織中乳酸化顯著增加,主要定位于mGCs。質(zhì)譜分析鑒定出組蛋白H4第5位賴氨酸(H4K5)的乳酸化,免疫組織化學(xué)染色顯示H4K5la在顆粒細(xì)胞中特異性富集,而卵母細(xì)胞和膜細(xì)胞信號極弱。這些發(fā)現(xiàn)表明乳酸衍生的H4K5la可能作為FSH誘導(dǎo)的mGCs線粒體生物發(fā)生的表觀遺傳介質(zhì)。
2.2 抑制乳酸產(chǎn)生/H4K5la抑制FSH誘導(dǎo)的顆粒細(xì)胞線粒體生物發(fā)生
為確定FSH是否通過乳酸/組蛋白乳酸化途徑促進(jìn)線粒體生物發(fā)生,研究人員在KGN細(xì)胞中測量了FSH處理后的細(xì)胞外酸化率(ECAR)。FSH顯著增加ECAR,表明糖酵解活性增強(qiáng)。為調(diào)節(jié)細(xì)胞內(nèi)組蛋白乳酸化,研究人員采用兩種互補(bǔ)方法:(a)使用2-脫氧-D-葡萄糖(2-DG)和草氨酸鹽藥理學(xué)抑制糖酵解通量以抑制乳酸產(chǎn)生;(b)siRNA介導(dǎo)的乳酸脫氫酶A(LDHA)和LDHB敲低。兩種抑制劑處理均有效降低mGCs中細(xì)胞內(nèi)乳酸水平,并顯著降低FSH刺激的mGCs和KGN細(xì)胞中的全局蛋白乳酸化和H4K5la水平。免疫熒光顯示2-DG和草氨酸鹽有效抑制FSH刺激的KGN細(xì)胞中H4K5la。組蛋白乳酸化降低顯著損害了FSH誘導(dǎo)的顆粒細(xì)胞線粒體生物發(fā)生,表現(xiàn)為線粒體DNA拷貝數(shù)、TOM20蛋白豐度和MitoTracker Green熒光減少。考慮到2-DG和草氨酸鹽的潛在脫靶效應(yīng),研究人員通過遺傳耗竭LDHA/B來特異性調(diào)節(jié)乳酸依賴性組蛋白乳酸化。雙重LDHA/B沉默顯著降低細(xì)胞內(nèi)乳酸生成、全局蛋白乳酸化和H4K5la。免疫熒光分析證實LDHA/B敲低后FSH刺激的H4K5la受損。相應(yīng)干預(yù)抑制了FSH誘導(dǎo)的線粒體生物發(fā)生,表現(xiàn)為線粒體DNA拷貝數(shù)減少、TOM20表達(dá)降低和MitoTracker Green染色減弱。為評估乳酸補(bǔ)充是否能挽救這一表型,研究人員用乳酸鈉處理LDHA/B耗竭的KGN細(xì)胞。外源性乳酸顯著恢復(fù)了FSH刺激細(xì)胞中的H4K5la和線粒體生物發(fā)生。此外,F(xiàn)SH處理顯著增強(qiáng)KGN細(xì)胞的氧消耗率(OCR),而抑制乳酸產(chǎn)生減弱了這一效應(yīng),乳酸補(bǔ)充wan全挽救了OCR。類似地,LDHA/B敲低降低了FSH刺激的ATP產(chǎn)生,乳酸鈉可恢復(fù)。使用JC-1熒光探針實時監(jiān)測線粒體膜電位(ΔΨm)發(fā)現(xiàn),F(xiàn)SH處理顯著增加紅色熒光(J-聚集體,表示超極化)同時降低綠色熒光(JC-1單體,表示去極化)。重要的是,藥理學(xué)抑制乳酸產(chǎn)生顯著減弱FSH誘導(dǎo)的ΔΨm升高,而乳酸鈉補(bǔ)充wan全挽救了這一超極化效應(yīng)。為確定FSH是否通過抑制線粒體自噬增強(qiáng)線粒體含量,研究人員評估了線粒體自噬標(biāo)志物。在FSH處理的KGN細(xì)胞中,PINK1/Parkin和LC3-II表達(dá)增加,而p62表達(dá)降低,與激活的線粒體自噬通量一致。FSH處理后線粒體-溶酶體共定位也增加。這些結(jié)果共同表明FSH通過乳酸/組蛋白乳酸化途徑而非抑制線粒體自噬促進(jìn)顆粒細(xì)胞線粒體生物發(fā)生。
2.3 FSH通過P300/CBP催化的H4K5la促進(jìn)線粒體生物發(fā)生
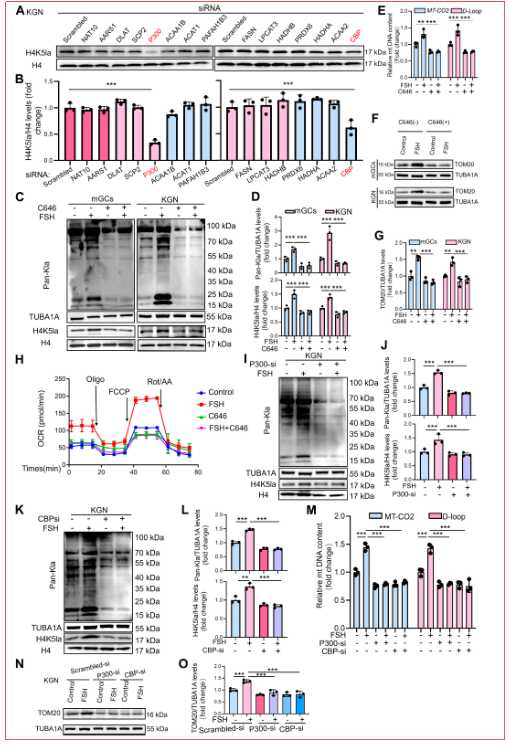
為鑒定負(fù)責(zé)H4K5la的酶,研究人員使用siRNA方法在KGN細(xì)胞中沉默候選酰基轉(zhuǎn)移酶。P300及其同源物CBP的敲低降低了H4K5la,鑒定P300/CBP為與該乳酸化特征相關(guān)的關(guān)鍵乳酸轉(zhuǎn)移酶。研究人員接下來使用C646(特異性P300/CBP拮抗劑)抑制該復(fù)合物。如預(yù)期,C646處理有效抑制FSH處理的mGCs和KGN細(xì)胞中的全局乳酸化和H4K5la。由于P300/CBP也可通過作為寫入蛋白催化乙酰化,研究人員評估了FSH處理的KGN細(xì)胞中的乙酰化水平。然而未觀察到顯著變化,且C646未消除全局乙酰化,表明P300/CBP不是wei一的乙酰轉(zhuǎn)移酶或其作用在這些細(xì)胞中被補(bǔ)償。接下來研究人員檢查了P300/CBP抑制后的線粒體生物發(fā)生。P300/CBP抑制消除了FSH誘導(dǎo)的線粒體DNA含量增加和TOM20表達(dá)。MitoTracker染色證實C646阻斷FSH刺激的線粒體生物發(fā)生。與此一致,C646顯著減弱FSH刺激的OCR和線粒體膜電位。為解決C646可能產(chǎn)生超出乳酸化抑制的脫靶效應(yīng)的擔(dān)憂,研究人員進(jìn)行了P300和CBP的單獨沉默。類似地,酶缺失顯著降低全局和H4K5乳酸化,并相應(yīng)減少KGN細(xì)胞中FSH誘導(dǎo)的線粒體生物發(fā)生,表現(xiàn)為線粒體DNA拷貝數(shù)、TOM20表達(dá)和MitoTracker Green熒光降低。P300/CBP敲低還抑制FSH誘導(dǎo)的線粒體膜電位升高。這些數(shù)據(jù)共同證明P300/CBP介導(dǎo)的H4K5la促進(jìn)FSH驅(qū)動的顆粒細(xì)胞線粒體生物發(fā)生。
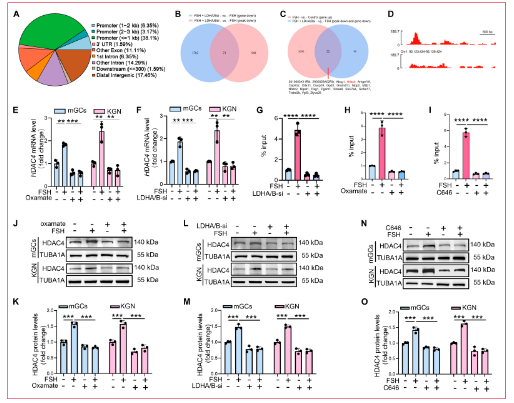
2.4 鑒定H4K5la調(diào)控的下游靶基因
為評估組蛋白乳酸化對FSH激活顆粒細(xì)胞中基因表達(dá)的影響,研究人員使用H4K5la特異性抗體進(jìn)行CUT&Tag分析。質(zhì)量控制分析證實樣本間高相關(guān)性和適當(dāng)?shù)腄NA片段大小分布。H4K5la峰主要富集于啟動子區(qū)域。GO分析顯示H4K5la靶基因在mGCs中強(qiáng)烈富集于代謝調(diào)控,而KEGG通路注釋同樣顯示在代謝網(wǎng)絡(luò)中富集,表明組蛋白乳酸化有助于線粒體生物發(fā)生的基礎(chǔ)代謝基因表達(dá)調(diào)控。為檢查功能后果,研究人員對LDHA/LDHB敲低后的FSH處理mGCs進(jìn)行RNA測序。差異表達(dá)分析顯示與FSH處理對照相比,F(xiàn)SH + LDHA/B-si組中有1,773個下調(diào)基因和1,335個上調(diào)基因。下調(diào)基因的KEGG富集指向核心代謝過程,表明乳酸依賴性乳酸化支持線粒體生物發(fā)生的基礎(chǔ)代謝基因表達(dá)。通過整合CUT&Tag數(shù)據(jù)(下調(diào)的H4K5la峰)與FSH處理GCs ± LDHA/B敲低的RNA-seq數(shù)據(jù)(下調(diào)基因),研究人員鑒定出71個H4K5la靶基因,其轉(zhuǎn)錄在LDHA/B耗竭后顯著降低。為進(jìn)一步將FSH誘導(dǎo)的H4K5la與線粒體生物發(fā)生聯(lián)系起來,研究人員分析了FSH處理mGCs的RNA-seq譜,鑒定出2,003個下調(diào)和1,718個上調(diào)基因。KEGG分析顯示上調(diào)轉(zhuǎn)錄本與線粒體生物發(fā)生通路密切相關(guān)。將這些基因與71個差異表達(dá)基因交叉參考,鑒定出22個被FSH顯著誘導(dǎo)且在LDHA/LDHB敲低后顯著下調(diào)的H4K5la靶基因。在這些基因中,HDAC4(一種作為乙酰化而非乳酸化擦除器的HDAC II類去乙酰化酶)在其啟動子處顯示強(qiáng)烈的H4K5la富集。與此發(fā)現(xiàn)一致,qRT-PCR證實FSH刺激的mGCs和KGN細(xì)胞中草氨酸鹽處理或LDHA/LDHB敲低后HDAC4 mRNA表達(dá)降低。ChIP-qPCR進(jìn)一步驗證了FSH刺激下HDAC4啟動子處的H4K5la富集,該富集被草氨酸鹽、LDHA/B沉默或使用C646抑制乳酸化寫入蛋白P300所減弱。免疫印跡顯示草氨酸鹽、LDHA/LDHB敲低或C646處理后HDAC4蛋白降低。免疫熒光染色證實抑制乳酸產(chǎn)生和H4K5la降低HDAC4蛋白表達(dá),HDAC4主要定位于細(xì)胞核。這些結(jié)果共同確立FSH誘導(dǎo)的H4K5la驅(qū)動顆粒細(xì)胞中HDAC4的轉(zhuǎn)錄上調(diào)。
2.5 HDAC4促進(jìn)FSH誘導(dǎo)的線粒體生物發(fā)生
在確立H4K5la調(diào)控HDAC4表達(dá)后,研究人員接下來研究了HDAC4在FSH處理顆粒細(xì)胞中的功能作用。使用LMK-235(特異性IIa類去乙酰化酶家族拮抗劑,據(jù)報道可在轉(zhuǎn)錄和翻譯水平抑制HDAC4表達(dá))處理顆粒細(xì)胞。如預(yù)期,LMK-235處理后KGN細(xì)胞中HDAC4表達(dá)以濃度依賴性方式降低。基于這些結(jié)果,選擇15 μM LMK-235作為后續(xù)實驗的標(biāo)準(zhǔn)濃度。通過qRT-PCR定量線粒體DNA含量、線粒體標(biāo)志物TOM20免疫印跡和MitoTracker Green染色線粒體證明,HDAC4的藥理學(xué)抑制顯著減弱FSH誘導(dǎo)的mGCs和KGN細(xì)胞線粒體生物發(fā)生。為小化LMK-235與HDAC4抑制無關(guān)的潛在脫靶效應(yīng),研究人員隨后采用siRNA介導(dǎo)的方法特異性沉默HDAC4。初步評估了四種靶向HDAC4的siRNA的基因沉默效率和特異性。FSH刺激下HDAC4敲低顯著損害線粒體生物發(fā)生,表現(xiàn)為線粒體DNA拷貝數(shù)減少、TOM20蛋白水平降低和MitoTracker Green熒光減弱。與此一致,HDAC4耗竭抑制KGN細(xì)胞中FSH誘導(dǎo)的OCR和ΔΨm增加。這些結(jié)果牢固確立HDAC4作為顆粒細(xì)胞響應(yīng)FSH的線粒體生物發(fā)生關(guān)鍵調(diào)節(jié)因子。

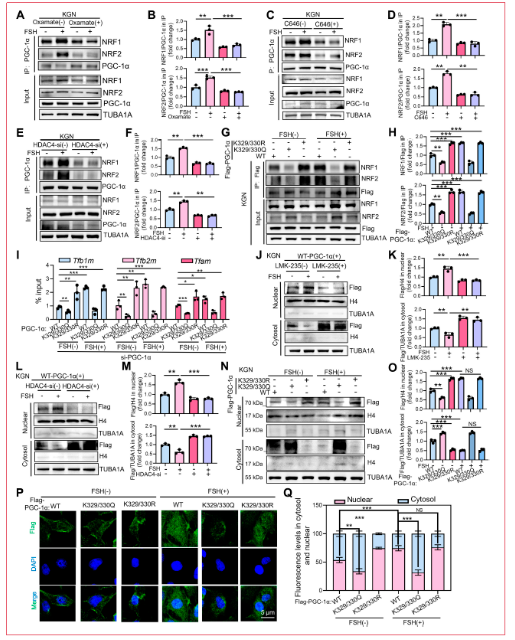
2.6 HDAC4通過位點特異性去乙酰化調(diào)節(jié)PGC-1α活性以促進(jìn)線粒體生物發(fā)生
PGC-1α是代謝途徑的既定主調(diào)節(jié)因子。先前研究表明HDAC4直接靶向PGC-1α并在賴氨酸殘基K329和K330去乙酰化以調(diào)節(jié)其活性,但這些位點的乙酰化是否影響線粒體生物發(fā)生仍未知。為解決這個問題,研究人員檢查了HDAC4介導(dǎo)的PGC-1α賴氨酸去乙酰化在線粒體調(diào)節(jié)中的作用。跨物種序列比對顯示PGC-1α K329/K330的強(qiáng)烈進(jìn)化保守性,突出其潛在功能重要性。研究人員首先評估了FSH刺激的KGN細(xì)胞中PGC-1α乙酰化,觀察到FSH處理后乙酰化顯著降低,P300/CBP抑制劑C646逆轉(zhuǎn)了這一效應(yīng)。類似地,HDAC4敲低增加了FSH處理細(xì)胞中PGC-1α乙酰化。接下來研究人員將K329/330殘基單獨突變?yōu)椴豢梢阴;木玜n酸(R)殘基或可乙酰化的谷an酰胺(Q)殘基,并檢查每個突變體暴露于FSH時的乙酰化水平。KGN細(xì)胞通過脂質(zhì)體轉(zhuǎn)染表達(dá)野生型(WT)Flag-PGC-1α或其乙酰化模擬(K329/330Q)和不可乙酰化(K329/330R)變體。為確保檢測直接乙酰化,免疫沉淀復(fù)合物用SDS解離后用泛乙酰賴氨酸抗體免疫印跡。K329/330R突變幾乎消除了FSH誘導(dǎo)的PGC-1α乙酰化,證實這些位點代表主要乙酰化位點。為繪制與PGC-1α相關(guān)的乙酰轉(zhuǎn)移酶,研究人員在NIH/3T3細(xì)胞中進(jìn)行IP后質(zhì)譜分析。這種方法定義了PGC-1α相互作用組并鑒定出1,472個推定相互作用蛋白,包括其乙酰化的潛在介質(zhì)。GO富集分析突出顯示這些相互作用因子與線粒體生物發(fā)生通路的強(qiáng)烈關(guān)聯(lián)。基于功能注釋優(yōu)先篩選出11個候選乙酰轉(zhuǎn)移酶。靶向siRNA篩選鑒定DLAT和ACAT1為PGC-1α翻譯后修飾的關(guān)鍵調(diào)節(jié)因子,其耗竭顯著降低PGC-1α乙酰化。后續(xù)co-IP實驗證實兩種酶與PGC-1α相互作用,這些相互作用被FSH刺激動態(tài)破壞。研究人員隨后評估PGC-1α乙酰化是否影響FSH誘導(dǎo)的線粒體生物發(fā)生。KGN細(xì)胞在siRNA介導(dǎo)的內(nèi)源性PGC-1α耗竭后轉(zhuǎn)染W(wǎng)T PGC-1α或K329/330R和K329/330Q突變體。K329/330R突變體(去乙酰化模擬)顯著增強(qiáng)線粒體生物發(fā)生,而乙酰化模擬K329/330Q突變體損害線粒體生物發(fā)生,表現(xiàn)為(a)線粒體DNA拷貝數(shù)減少,(b)TOM20水平降低,和(c)MitoTracker Green染色減少。相應(yīng)地,K329/330R突變體升高線粒體OCR、ATP水平和ΔΨm,而K329/330Q突變體產(chǎn)生相反效應(yīng)。這些發(fā)現(xiàn)共同證明FSH通過HDAC4介導(dǎo)的PGC-1α K329/330去乙酰化促進(jìn)線粒體生物發(fā)生。

2.7 PGC-1α去乙酰化招募NRF1/2以驅(qū)動線粒體生物發(fā)生
PGC-1α作為線粒體生物發(fā)生的主調(diào)節(jié)因子,協(xié)同轉(zhuǎn)錄因子如NRF1和GABPA(NRF2)激活線粒體功能所需的核基因。為確定K329/330乙酰化如何影響PGC-1α活性,研究人員通過co-IP檢查FSH依賴性變化。FSH刺激顯著增強(qiáng)PGC-1α與NRF1/NRF2的相互作用,而破壞乳酸依賴性H4K5la消除這一效應(yīng)。類似地,暴露于P300/CBP抑制劑C646或HDAC4耗竭顯著減弱KGN細(xì)胞中FSH誘導(dǎo)的PGC-1α-NRF1/2復(fù)合物形成。為評估K329/330乙酰化的功能意義,研究人員在KGN細(xì)胞中表達(dá)Flag標(biāo)記的PGC-1α變體(WT、乙酰化缺陷K329/330R或乙酰化模擬K329/330Q)。K329/330R突變增強(qiáng)FSH刺激下PGC-1α與NRF1/NRF2的結(jié)合,而K329/330Q變體顯著損害這些相互作用。與此一致,ChIP-qPCR顯示FSH處理細(xì)胞中K329/330R突變體對NRF1/NRF2靶基因(TFAM、TFB1M、TFB2M)啟動子的占據(jù)增加,而K329/330Q突變體染色質(zhì)結(jié)合顯著降低。這些結(jié)果共同表明PGC-1α在K329/330的去乙酰化促進(jìn)其招募至NRF1/2并驅(qū)動響應(yīng)FSH的線粒體基因程序激活。PGC-1α必須從細(xì)胞質(zhì)轉(zhuǎn)位至細(xì)胞核以發(fā)揮轉(zhuǎn)錄輔激活因子功能。為確定乙酰化是否調(diào)節(jié)其亞細(xì)胞分布,研究人員用Flag標(biāo)記的WT PGC-1α轉(zhuǎn)染KGN細(xì)胞并進(jìn)行核-胞質(zhì)分級分離。FSH處理誘導(dǎo)PGC-1α強(qiáng)烈的核定位,而LMK-235抑制HDAC4或HDAC4特異性siRNA減弱這一核積累。為進(jìn)一步探究K329/330乙酰化的作用,研究人員表達(dá)WT、K329/330R或K329/330Q PGC-1α構(gòu)建體后進(jìn)行亞細(xì)胞分級分離。K329/330R突變體增強(qiáng)FSH刺激的PGC-1α核進(jìn)入,而K329/330Q突變體減弱核定位。免疫熒光染色證實這些結(jié)果,顯示FSH處理細(xì)胞中K329/330R變體的胞質(zhì)-核轉(zhuǎn)位增加和K329/330Q突變體的核-胞質(zhì)重新分布增強(qiáng)。這些發(fā)現(xiàn)表明FSH降低PGC-1α乙酰化,促進(jìn)其核進(jìn)入并增強(qiáng)線粒體生物發(fā)生。
2.8 抑制H4K5la/HDAC4/PGC-1α通路損害體內(nèi)FSH誘導(dǎo)的線粒體生物發(fā)生和卵泡發(fā)育
為評估H4K5la/HDAC4/PGC-1α通路的生理相關(guān)性,接受FSH的小鼠用靶向該軸關(guān)鍵組分的抑制劑處理:C646(P300/CBP乳酸化抑制劑)、LMK-235(HDAC4拮抗劑)或SR-18292(PGC-1α乙酰化激活劑)。C646處理消除了卵巢顆粒細(xì)胞中FSH誘導(dǎo)的H4K5la,并伴隨抑制HDAC4表達(dá)。與HDAC4介導(dǎo)的PGC-1α去乙酰化一致,通路阻斷增加PGC-1α乙酰化并破壞PGC-1α-NRF1/NRF2相互作用。重要的是,該通路阻斷顯著損害FSH誘導(dǎo)的線粒體生物發(fā)生,表現(xiàn)為線粒體基因表達(dá)降低、mtDNA拷貝數(shù)減少和TOM20蛋白水平降低。線粒體缺陷伴隨雌二醇(E2)產(chǎn)生減少和顆粒細(xì)胞增殖降低。在器官水平,抑制劑處理的小鼠表現(xiàn)出卵巢變小、卵巢重量降低、卵泡發(fā)育停滯和竇卵泡形成減少。這些發(fā)現(xiàn)共同證明H4K5la/HDAC4/PGC-1α軸對FSH介導(dǎo)的卵泡發(fā)育至關(guān)重要,通過支持線粒體代謝適應(yīng)。
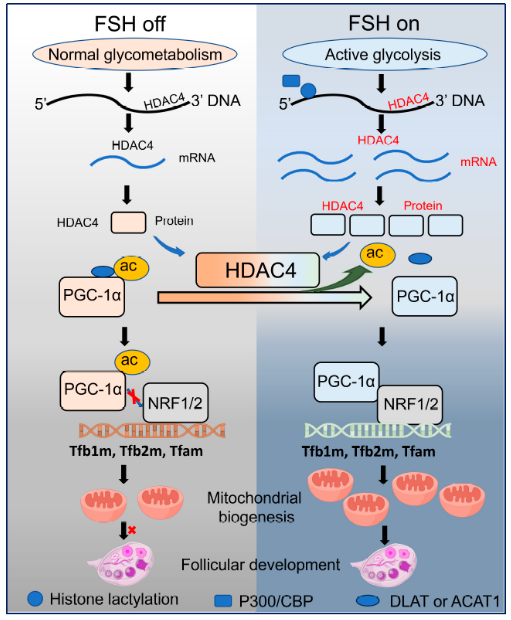
n 研究結(jié)論
本研究揭示了一個先前未表征的FSH依賴性代謝-表觀遺傳通路,將糖酵解與線粒體生物發(fā)生聯(lián)系起來。FSH刺激的糖酵解增加細(xì)胞內(nèi)乳酸,通過P300/CBP介導(dǎo)的H4K5組蛋白乳酸化(H4K5la),該修飾富集于代謝基因啟動子并直接激活HDAC4轉(zhuǎn)錄。HDAC4介導(dǎo)的PGC-1α在K329/K330去乙酰化促進(jìn)NRF1/2招募和線粒體DNA復(fù)制及線粒體生物發(fā)生必需基因的轉(zhuǎn)錄。該機(jī)制可能代表代謝通量控制細(xì)胞器生物發(fā)生表觀遺傳調(diào)控的更廣泛原理,為理解FSH依賴性生殖生理提供了機(jī)制見解,并突出了生殖障礙代謝功能障礙的潛在治liao機(jī)會。
Wu G, Chen M, Li C, Wei M, Pan Y, He T, Liu Z, Li H, Zhang C, Zhang JQ, Sheng Y, Liu Y, Liu H, Shen M. Histone Lactylation Couples FSH-Driven Lactate Metabolism to Mitochondrial Biogenesis by Enhancing HDAC4-Mediated Deacetylation of PGC-1α in Granulosa Cells. Research (Wash D C). 2026 Jan 15;9:1045. doi: 10.34133/research.1045. PMID: 41551912; PMCID: PMC12804605.

電話
微信掃一掃